Full Bio
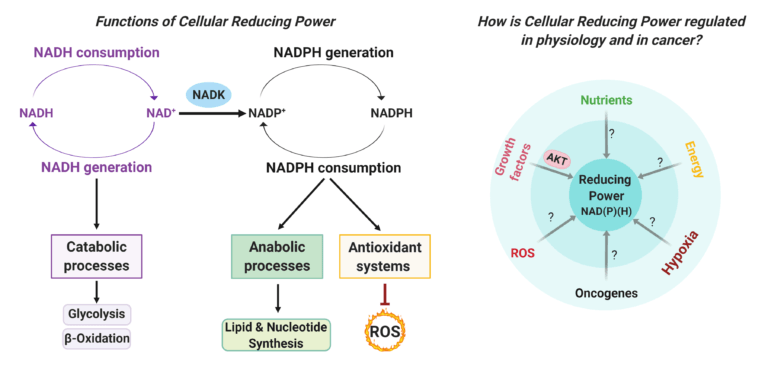
Cellular reducing power, which is stored in the pyridine nucleotide cofactors NAD(H) and its phosphorylated form NADP(H), drives more than 500 biochemical reactions, including ATP generation, biosynthesis of macromolecules, and redox homeostasis. Dysregulation of cellular reducing power has been linked to many diseases, including cancer. However, the underlying mechanisms of this dysregulation are not well understood.
Recently, we identified the oncogenic PI3K-Akt pathway as a critical regulator of the cellular reducing power through direct stimulation of NAD+ kinase (NADK) (Science 363, 1088-1092, 2019). NADK catalyzes the phosphorylation of NAD+ to NADP+, which is then used to generate NADPH. Despite its importance, the mechanistic regulation of NADK and its cellular and metabolic functions are grossly understudied. Furthermore, the role of NADK in normal tissues and in cancer cells remains poorly defined.
Our laboratory is interested in identifying new regulators of NADK function and cellular reducing power in cancer. We employ quantitative metabolomics, biochemistry, proteomics, and mouse models to obtain a holistic understanding of the regulation of cellular reducing power in physiology and disease. Elucidating the pathways that control cellular redox in cancer could reveal new liabilities that can be exploited for cancer therapies.
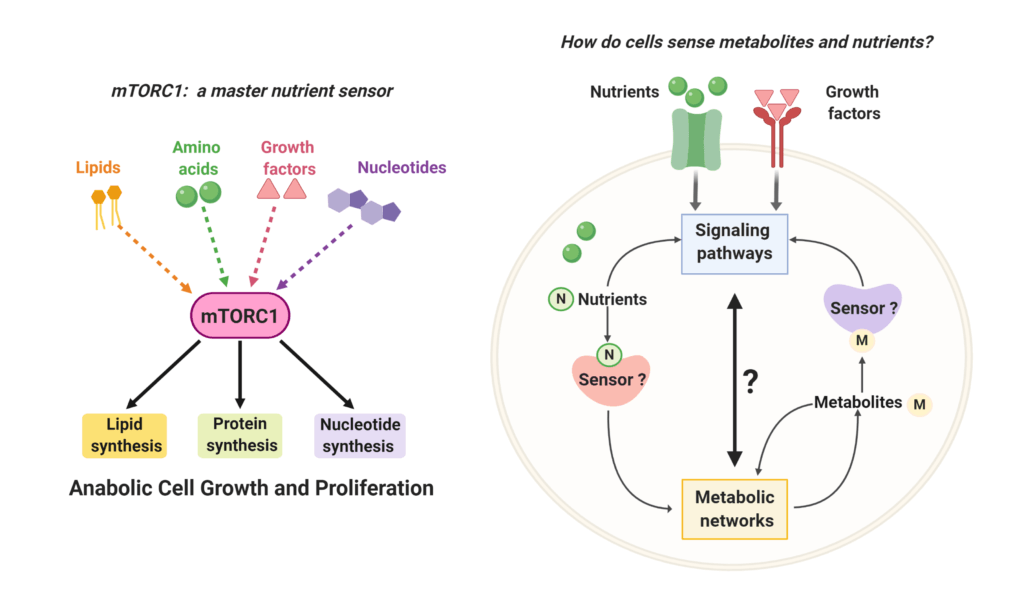
The ability to sense metabolites and nutrients is one of the most fundamental properties of all organisms and cells that has allowed for the adaptation, survival, and thriving of species in different environmental and cellular conditions. Metabolite-sensing systems allow for proper communication and calibration of cellular signaling and metabolic status. A classic example is the ability of the protein kinase mTORC1 to sense and integrate signals from growth factors, cellular amino acid levels, and energy status to couple anabolic processes with nutrient and energy availability. In addition to the aforementioned inputs, we recently discovered that adenine nucleotides can also be sensed by mTORC1, but the key sensor(s) that relay this information remain unidentified (Cell Reports 21, 1331-1346, 2017).
Our laboratory is interested in studying how cells sense key metabolites and nutrients. We seek fundamental insights into how nutrient availability and metabolite sensing are coordinated with signaling networks for metabolic control, not only in physiological settings, but also in disease settings, where such coordination is lost.
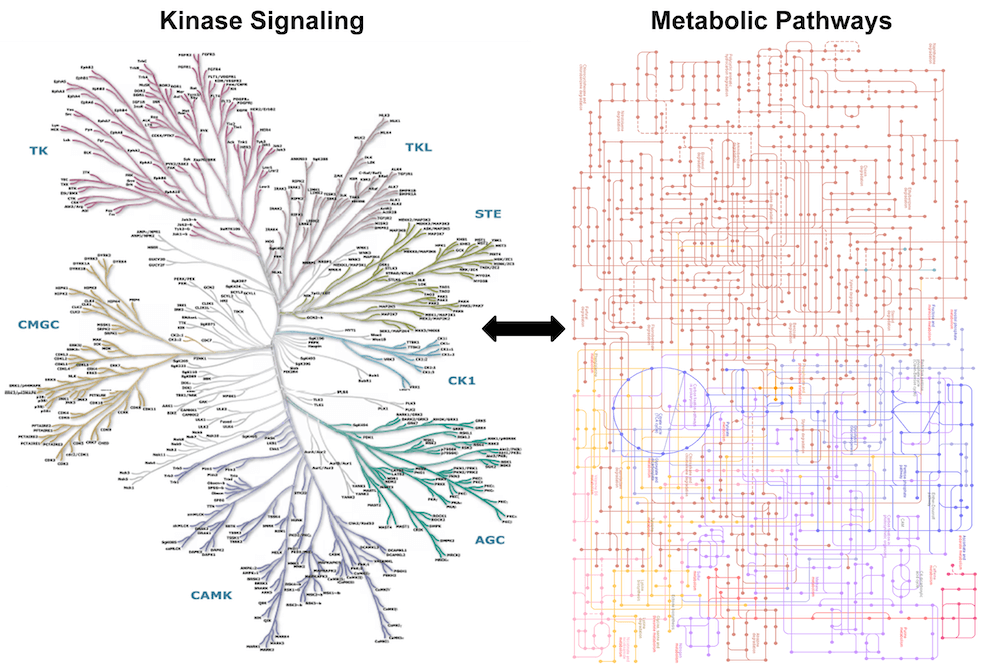
Rewiring of cellular metabolism is a hallmark of cancer, yet only a handful of metabolic enzymes are known to be mutated in cancer. While much work has been done in recent years to unravel the mechanisms leading to such metabolic alterations, we still have a limited understanding of the oncogenic events that reprogram metabolism in cancer cells.
Work in the past decade has revealed that signaling networks, which are frequently mutated in cancer, can recalibrate cellular metabolism to meet the anabolic demands of cancer cell proliferation (Nature Reviews Cancer 20, 74-88, 2020). Protein kinases are key regulators of signal transduction with vast roles in orchestrating cellular processes, including metabolism. However, out of the 518 kinases in the human kinome, only a small fraction have been implicated in regulating metabolism. Our lab is interested in identifying key kinases that control cancer metabolism. Discovering critical regulatory nodes at the intersection of the kinome and cellular metabolism has the potential to reveal new therapeutic targets.